
 18 апреля, 2018
18 апреля, 2018 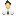 admin
admin 

Рис. 1. Представители «линий-сирот»: A — Ancyromonas (иллюстрация из Википедии), Б — Rigifila (иллюстрация из статьи A. Yabuki et al., 2013. Rigifila ramosa n. gen., n. sp., a filose apusozoan with a distinctive pellicle, is related to Micronuclearia), В — Mantamonas (иллюстрация из статьи E. Gluksman et al., 2011. The novel marine gliding zooflagellate genus Mantamonas (Mantamonadida ord. n.: Apusozoa)), Г — Collodictyon (иллюстрация с сайта dailymail.co.uk). Изображения в разных масштабах! На фото Б стрелки указывают на бактерий, захваченных длинными тонкими ветвящимися ложноножками ригифилы. На фото В стрелка указывает на передний жгутик
Новое молекулярно-филогенетическое исследование позволило выделить эволюционную ветвь эукариот, имеющую очень высокий ранг, но состоящую целиком из одноклеточных организмов: она включает малоизвестные группы коллодиктионид, мантамодад и ригифилид. При этом ее ранг значительно выше, чем, например, ранг ветви заднежгутиковых, в которую входят грибы вместе с многоклеточными животными (последняя находится внутри мегагруппы Amorphea). Если это открытие подтвердится, оно может потребовать серьезного пересмотра представлений о том, как устроено эволюционное древо эукариот в целом.
Одним из самых ярких достижений биологии начала XXI века стало создание совершенно новой системы эукариот — обладателей сложной клетки. Эукариоты — гигантская группа организмов, в которую среди прочих входят многоклеточные животные и зеленые растения. Область биологии, исследующая границы и родственные связи групп очень высокого ранга (таких, как животные и растения), называется мегасистематикой. Попросту говоря, мегасистематика — это «царствоведение» (В. А. Красильников, 2004. «Царствоведение», или мегасистематика). Напомним, что царством называется высшая систематическая единица живой природы; в традиционной классификации царствами считались животные, растения, грибы. Правда, в современной биологии термин «царство», как и названия других традиционных систематических рангов, понемногу выходит из употребления.
Сейчас выделяется от 5 до 8 групп эукариот, соответствующих главным эволюционным ветвям — они называются супергруппами. В свою очередь, супергруппы объединяются в совсем уж небольшое число мегагрупп, которых в последнее время насчитывают три.
Мегагруппа, к которой относимся мы сами, называется Amorphea или Unikonta. Кроме многоклеточных животных, в нее входят воротничковые жгутиконосцы, грибы, многочисленные голые и раковинные амебы, слизевики (точнее, миксомицеты) и еще несколько групп одноклеточных и колониальных организмов, малоизвестных широкой публике (см., например: Ядра мезомицетозоев делятся синхронно, как у зародышей животных, «Элементы», 05.06.2013). Название Amorphea буквально означает «бесформенные». Наша мегагруппа названа так потому, что многие ее одноклеточные представители склонны использовать для движения или для питания временные клеточные выросты — ложноножки, а клетка, регулярно выпускающая и втягивающая ложноножки, по определению не может иметь постоянной формы. Второе название этой мегагруппы — Unikonta — значит «одножгутиковые». Дело в том, что у тех одноклеточных представителей этой группы, которые передвигаются с помощью жгутиков (такие существа называются жгутиконосцами), жгутик действительно чаще всего один, в то время как у членов других мегагрупп эукариот их обычно два. Но это не абсолютный закон: обладатели двух жгутиков среди униконтов, особенно примитивных, тоже встречаются.
Самая разнообразная из трех мегагрупп эукариот — безусловно, мегагруппа Diaphoretickes, название которой по-гречески и значит «разнообразные». В эту мегагруппу входят всевозможные водоросли (бурые, золотистые, желто-зеленые, диатомовые, красные, зеленые, харовые), высшие растения, ложные грибы (оомицеты), споровики, инфузории, радиолярии, фораминиферы, — и этот список более чем далек от полноты. Неудивительно, что внутренняя система этой мегагруппы сложна, запутана и до сих пор полна нерешенных вопросов (см., например: Новооткрытый жгутиконосец проливает свет на раннюю эволюцию эукариот, «Элементы», 17.01.2018). Подавляющее большинство эукариот, имеющих хлоропласты и способных к фотосинтезу, относится именно к мегагруппе Diaphoretickes. Другое ее название — Corticata, что означает «имеющие кортекс» (в данном случае этим словом называется укрепленный внешний слой клетки). Развитие прочного кортекса в общем случае мешает питанию животного типа, основанному на активном захвате пищевых объектов, и лучше сочетается с питанием растительного типа — фотосинтезом. Но, во-первых, тут встречаются исключения (например, инфузории обошли это ограничение и остались хищниками, создав постоянный клеточный рот), а во-вторых, само понятие «кортекс», честно говоря, довольно расплывчато. Структуры, укрепляющие внешний слой клетки, бывают устроены очень по-разному и сочетаются с различным образом жизни, так что далеко не все кортикаты фотосинтезируют; среди них есть не только одноклеточные хищники, но и паразиты, и сапротрофы, питающиеся мертвыми остатками других организмов.
Третья мегагруппа эукариот называется Excavata. Ее название происходит от латинских слов ex — внешний и cava — полость, борозда (имеется в виду характерная для представителей этой мегагруппы ротовая бороздка — продольное углубление на той стороне клетки, которая условно считается брюшной). Мегагруппа Excavata состоит преимущественно из жгутиконосцев; колониальных форм в ней мало, а обладателей многоклеточности в строгом смысле слова нет вовсе. Именно к экскаватам относятся жгутиконосцы, вошедшие в школьные учебники зоологии — фотосинтезирующая эвглена и хищный бодо.
Итак, эволюционное древо эукариот распадается на три мегаствола: Amorphea (= Unikonta), Diaphoretickes (= Corticata) и Excavata. Примем во внимание, что это предельно беглый обзор; для обсуждения более дробных подразделений — даже такого ранга, как многоклеточные животные или высшие растения — в короткой статье нет места, ибо их слишком много. Но даже столь общая картина позволяет сделать несколько важных выводов.
Во-первых, рассмотренная нами система не имеет ничего общего с той системой живой природы, которая господствовала в учебниках на протяжении XX века. Например, большинство тех организмов, которые в классической биологии назывались «одноклеточными животными», на самом деле относится или к мегагруппе Diaphoretickes, или к мегагруппе Excavata. Это означает, что они не более родственны животным, чем, скажем, какие-нибудь зеленые растения. С другой стороны, довольно близкими (по меркам мегасистемы) родственниками многоклеточных животных оказались грибы — раньше сближать эти группы было совершенно не принято.
Во-вторых, колоссальное место в современной системе эукариот занимают организмы, которые, в отличие от всем известных животных и растений, отнюдь не являются многоклеточными (во всяком случае, истинно многоклеточными). Еще с XIX века их принято собирательно называть протистами, то есть буквально «простейшими». К протистам в самом широком понимании этого термина можно при желании отнести всех эукариот, кроме многоклеточных животных и высших растений (J. O. Corliss, 1989. Protistan diversity and origins of multicellular/multitissued organisms). Гораздо чаще протистами называют все же только одноклеточных эукариот (или колониальных — при условии, что колонии состоят из однотипных клеток, способных в принципе жить отдельно). Но таких все равно очень много, и из них могут состоять целые супергруппы. По сути, все основание эволюционного древа эукариот слагается из протистов. Группы, состоящие целиком из многоклеточных организмов, образуют на этом древе лишь редкие «выбросы». Например, ветвей такого порядка, как многоклеточные животные, на эволюционном древе эукариот можно насчитать несколько сотен, и в большинстве из них истинная многоклеточность никогда не возникала (F. Burki, 2014. The eukaryotic tree of life from a global phylogenomic perspective).
В-третьих, примитивными членами всех без исключения мегагрупп и супергрупп эукариот оказались одноклеточные протисты, передвигающиеся с помощью жгутиков, то есть уже не раз упоминавшиеся жгутиконосцы. Многие эволюционные ветви достаточно высокого ранга состоят из жгутиконосцев вообще целиком. Ни одна другая жизненная форма не имеет на эволюционном древе эукариот такого всеохватывающего распространения. Это — явное свидетельство того, что общий предок всех современных эукариот тоже был жгутиконосцем (скорее всего, имевшим 2 жгутика).
Но в любом случае система эукариот не исчерпывается этим простым описанием. Дело в том, что существует не столь уж малое количество эволюционных линий протистов, которые, судя по молекулярным данным, не входят ни в одну известную супергруппу, и даже их принадлежность к мегагруппам зачастую остается под вопросом. В последнее время их называют «линиями-сиротами» («orphan» lineages). В подавляющем большинстве это — конечно же — жгутиконосцы. К тому же многие из «линий-сирот» явно расположены очень близко к общему корню эукариотного древа. Именно поэтому уточнение их родственных связей может оказаться крайне важным для понимания эволюции эукариот в целом.
Новая супергруппа
Недавно вышла очередная статья, уточняющая структуру эволюционного древа эукариот на основе молекулярно-биологических данных. Список авторов этой статьи замыкают известные канадские биологи Аластер Симпсон (Alastair Simpson) и Эндрю Роджер (Andrew Roger), которые уже много лет решают молекулярными методами проблемы мегасистематики эукариот; их мнение служит хорошим показателем текущего положения дел в этой области (см., например: A. G. B. Simpson, A. J. Roger, 2004. The real ‘kingdoms’ of eukaryotes).
На этот раз в центре внимания Роджера, Симпсона и их коллег оказались четыре группы одноклеточных организмов, относящихся к «линиям-сиротам» (рис. 1).
- Коллодиктион (см. Collodictyon) — пресноводный жгутиконосец с 4 жгутиками и небольшими ложноножками, склонный к питанию крупной по своим меркам добычей. Вместе с родственным жгутиконосцем, имеющим два жгутика (см. Diphylleia), он выделяется в семейство коллодиктионид (см. Collodictyonidae).
- Анкиромонады (см. Ancyromonas) — жгутиконосцы, живущие в воде или почве. Для них характерно наличие двух жгутиков, один из которых направлен вперед, а другой назад.
- Мантамонады (см. Mantamonas) — морские жгутиконосцы с двумя жгутиками и небольшими ложноножками, живущие в прибрежных донных осадках.
- Ригифилиды (см. Rigifila) — пресноводные протисты без жгутиков, зато с ложноножками особого типа — длинными и тонкими (они называются филоподиями).
Роджер, Симпсон и их коллеги построили общее древо эукариот, включающее перечисленные «линии-сироты» вместе много с кем еще и основанное на солидном наборе данных — программа обработала выравненные последовательности 351 белка (97 000 аминокислотных остатков). Результаты получились интересные.
Прежде всего, оказалось, что три из четырех названных «линий-сирот» — коллодиктиониды, мантамонады и ригифилиды — образуют на древе единую компактную эволюционную ветвь, вполне заслуживающую статуса супергруппы. Гипотеза о единстве этой ветви обоснована молекулярными данными достаточно хорошо, чтобы новая супергруппа получила предварительное название — CRuM (сокращение от названий входящих в нее семейств). Это — совершенно новое объединение. Раньше коллодиктионид, мантамонад и ригифилид куда только не помещали, но сейчас авторы готовы настаивать, что решили задачу верно: их новая гипотеза имеет очень высокую поддержку.
На общем древе ветвь CRuM примыкает к основанию ветви Amorphea (= Unikonta), образуя сестринскую группу по отношению к ней (рис. 2).


Рис. 2. Новая версия эволюционного древа эукариот. Иллюстрация из обсуждаемой статьи в Genome Biology and Evolution
Третья упомянутая нами «линия-сирота» — анкиромонады — пока осталась в одиночестве. На общем древе ветвь анкиромонад находится недалеко от основания ветви Amorphea (= Unikonta), но все же дальше от него, чем CRuM. Кстати, примерно такое же положение на общем древе занимает и еще одна маленькая группа пресноводных жгутиконосцев — малавимонады (см. Malawimonas). Однако в единую группу анкиромонады и малавимонады не сливаются (во всяком случае, данные на этот счет неоднозначны).
Получен и еще один результат, не вынесенный в заголовок обсуждаемой статьи, но тем не менее очень интересный. Симпсон, Роджер и их коллеги показали, что мегагруппа Excavata скорее всего на самом деле не является единой ветвью. Согласно полученным данным, эта группа, почти целиком состоящая из жгутиконосцев, распадается на две эволюционных линии, одна из которых несколько примитивнее другой. Первая эволюционная линия «экскаватных» жгутиконосцев называется метамонадами (Metamonada), вторая — дискобами (Discoba); именно к дискобам относятся, например, упомянутые выше эвглена и бодо. Если все это верно, то получается, что Excavata — не ветвь, а эволюционный уровень. Этот вывод нельзя назвать потрясающей сенсацией, потому что близкие к нему гипотезы высказывались и раньше, причем достаточно регулярно (см., например: T. Cavalier-Smith et al., 2014. Multigene eukaryote phylogeny reveals the likely protozoan ancestors of opisthokonts (animals, fungi, choanozoans) and Amoebozoa). Тем не менее, любые новые данные здесь достойны пристального внимания. Речь ведь идет о том, как устроено все древо эукариот целиком и с чего начиналась их эволюция — не больше и не меньше.
Почему сие важно?
Знаменитый генетик Николай Владимирович Тимофеев-Ресовский имел обыкновение на научных семинарах, выслушав докладчика, громко спрашивать: «Почему сие важно в-пятых?» Этот заимствованный из русской классики вопрос призывал участников как бы отступить на шаг, чтобы увидеть проблему в целом и понять точно — какое место в общей картине занимают обнаруженные факты.
При обсуждении научных новостей эта постановка вопроса тоже бывает полезна. Действительно, какое значение для нашего мировоззрения может иметь очередная перетасовка запутанных родственных связей между мало кому известными эукариотными микроорганизмами, живущими где-нибудь в далеких морях или болотах? И на какие детали тут стоит обращать внимание, а на какие нет?
Чтобы разобраться в этом, начнем с фактов, которые мы знаем более-менее твердо. Все современные эукариоты имеют единое происхождение: они произошли от одного предка, а не от нескольких эволюционных линий. Никаких эпизодов «параллельной эукариотизации», насколько можно судить, не было — по крайней мере, на тех эволюционных уровнях, которые доступны сейчас для изучения (см. Теория симбиогенеза 50 лет спустя: параллельной эукариотизации, скорее всего, не было, «Элементы», 22.11.2017). Не вызывает сомнений, что общий предок всех современных эукариот был «полностью оперившимся», типичным по нашим меркам эукариотом, у которого уже было полностью сформированное ядро, цитоскелет, эндоплазматическая сеть, а также митохондрии и жгутики. Облик этого предполагаемого предка в большой степени сохранили коллодиктиониды, мантамонады, анкиромонады, малавимонады (и некоторые другие жгутиконосцы тоже). Не случайно все они, в общем, достаточно сильно похожи друг на друга. Итак, есть серьезные основания считать, что общий предок современных эукариот был гетеротрофным жгутиконосцем, который вел придонный образ жизни, питался в основном бактериями, имел два жгутика и в придачу к ним небольшие ложноножки, служившие на том этапе больше для питания, чем для движения.
В каком же порядке от этого общего предка расходились эволюционные ветви? Тут начинаются сложности. Почти все основные супергруппы эукариот молекулярная систематика (стоило ей только возникнуть) выделила довольно легко, но вот установить их родственные отношения между собой оказалось очень трудно. Современная систематика, как правило, соблюдает методические ограничения кладистики, где по умолчанию предполагается, что эволюционное древо может ветвиться только дихотомически: в каждом узле каждая ветвь делится строго надвое. Поэтому молекулярные систематики видят свою задачу в сведении эволюционного древа к последовательности развилок надвое — дихотомий. И внутри эукариотных супергрупп это в целом удается. Проблемы начинаются при попытке раскрыть самую основу эукариотного древа — ту его часть, где «действующими лицами» становятся объединения супергрупп, мегагруппы и, конечно, «линии-сироты». Вот здесь сведение к набору дихотомий или не удается вовсе, или возникающие при этом гипотезы имеют недостаточную поддержку и легко ставятся под сомнение другими группами исследователей (см., например: D. He et al., 2014. An alternative root for the eukaryote tree of life). Почему?
Еще на рубеже XX и XXI веков французский биолог Эрве Филипп (Herve Philippe) предположил, что эта неопределенность имеет фундаментальную причину, связанную с особенностями ранней эволюции эукариот (см., например: H. Philippe et al., 2000. The new phylogeny of eukaryotes). Когда в истории жизни на Земле возникает что-то принципиально новое, это может сопровождаться кладогенезом — распадом маленькой предковой группы сразу на несколько эволюционных ветвей довольно высокого порядка. Кладогенез — вполне реальный способ эволюции. При этом свести его к последовательности дихотомий в общем случае принципиально невозможно, сколько ни старайся: что поделать, если группа-предок за короткое по эволюционным меркам время распалась не на две ветви, а на 3, 4, 5 или 6? Здесь кладистический подход просто обязан дать сбой. Эрве Филипп считал, что именно такая ситуация и наблюдается в начале эволюции эукариот (во всяком случае, их современных групп).
Не будем спешить с выводами. Даже если Филипп совершенно прав (что не доказано), это ни в коем случае не значит, что исследования, подобные тем, которые сейчас ведут Симпсон и Роджер, бесполезны. Совсем нет — эти исследования в любом случае дают новую, очень ценную информацию. Но вот помещение той или иной «линии-сироты» в ту или иную точку общего древа теперь может и не иметь решающего значения. Потому что биологический смысл складывающейся картины в любом случае уже приблизительно понятен — перестановки отдельных линий (которые вполне могут еще случиться) вряд ли его изменят. Вероятно, все современные эукариоты произошли от очень компактной группы предков, которые были жгутиконосцами, в целом похожими на современных анкиромонад или коллодиктионид. После серии быстрых расхождений (порядок которых, честно говоря, не столь важен) оформилось несколько эволюционных линий — например, на рис. 3 их семь, и порядок их расположения относительно друг друга там соответствует тому, что установили Симпсон и Роджер в обсуждаемой работе. Вероятно, это и есть картина кладогенеза.

Рис. 3. Сильно упрощенная версия предыдущего древа. Красный треугольник обозначает предполагаемый общий корень и указывает на общего предка всех современных эукариот
Остается выяснить, где же находится «корень» эукариотного древа? Это — очень непростой вопрос. Симпсон и Роджер принимают версию, согласно которой «корень» общего древа располагается между анкиромонадами и метамонадами, то есть между двумя группами жгутиконосцев (R. Derelle et al., 2015. Bacterial proteins pinpoint a single eukaryotic root). В некотором смысле можно сказать, что все эукариоты — жгутиконосцы. Просто некоторые из них сохранили эту жизненную форму до сих пор, а другие преобразовали ее, создав причудливые организмы амеб, инфузорий, грибов, растений, животных.
Источник: elementy.ru
 Опубликовано в рубрике
Опубликовано в рубрике