
 22 февраля, 2018
22 февраля, 2018 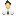 admin
admin 


Рис. 1. Из десяти энхансеров, управляющих работой генов, необходимых для развития конечностей, можно удалить любой, и это никак не повлияет на нормальное развитие конечности. Для каждого энхансера указано его название (например, mm1179), ген, чью работу он регулирует (например, Gli3), расстояние от энхансера до точки начала транскрипции; левая фотография в каждой паре показывает, в каких областях зачатка передней конечности (на 11-е сутки после оплодотворения) срабатывает данный энхансер, обеспечивая включение подконтрольного гена; правая фотография — передняя конечность позднего эмбриона (18-е сутки), у которого данный энхансер отсутствует в обеих хромосомах. Во всех случаях конечность получилась абсолютно нормальная. Изображение из обсуждаемой статьи в Nature
В геномах млекопитающих число регуляторных участков ДНК — энхансеров — на порядок превышает число белок-кодирующих генов. Смысл такого обилия регуляторов неясен. Эксперименты на генно-модифицированных мышах, которым при помощи CRISPR-Cas9 удаляли те или иные эволюционно консервативные (то есть, по-видимому, важные) энхансеры, показали, что функции многих энхансеров «избыточны»: эти регуляторы не дополняют, а дублируют друг друга. Дублирование повышает устойчивость развития, обеспечивая формирование нормального фенотипа невзирая на различные помехи.
Энхансеры (см. Enhancer) — регуляторные участки ДНК, к которым присоединяются специальные белки (факторы транскрипции). Каждый энхансер регулирует работу какого-нибудь белок-кодирующего гена, определяя, при каких условиях, в каких частях организма и на каких стадиях развития ген будет экспрессироваться. Многие новые признаки в ходе эволюции появляются за счет изменения энхансеров, а не самих белок-кодирующих генов (см., например: Генетические механизмы формирования сложных признаков постепенно проясняются, «Элементы», 14.04.2010). Чтобы выяснить функцию энхансера, то есть определяемый им пространственно-временной паттерн экспрессии, используют генно-инженерные эксперименты: вставляют в геном какой-нибудь «хорошо заметный» ген (например, ген флуоресцирующего белка), к которому приделан интересующий ученых энхансер, и смотрят, где и когда будет работать ген.
У одного и того же гена может быть несколько энхансеров с разными функциями. Эти регуляторы включают ген на разных стадиях развития и в разных тканях. Но иногда множественные энхансеры дублируют друг друга, определяя один и тот же паттерн экспрессии. Возникает вопрос: зачем нужно такое дублирование? В опытах на дрозофиле ранее было показано, что кажущаяся избыточность энхансеров может повышать устойчивость развития. Если что-то не заладится с одними регуляторными системами, другие их подстрахуют, и организм всё равно разовьется нормально. Таким образом, «лишние» энхансеры повышают надежность онтогенеза и обеспечивают формирование нормального фенотипа невзирая на помехи — внешние (средовые) или внутренние (например, мутационные) (см.: Избыточные регуляторы делают развитие эмбриона помехоустойчивым, «Элементы», 03.06.2010). Насколько широко такое подстраховывающее дублирование распространено в природе, еще предстоит выяснить.
У млекопитающих общее число энхансеров в геноме превышает число белок-кодирующих генов примерно на порядок. В чем смысл такого изобилия регуляторов, пока неясно. Чтобы в этом разобраться, хорошо бы для начала выяснить, какой тип «взаимоотношений» более типичен для энхансеров одного и того же гена — дополнение (когда разные энхансеры заставляют ген работать в разных местах в разное время) или дублирование. В первом случае число энхансеров, возможно, отражает прежде всего сложность строения организма, во втором — скорее «забуференность» и помехоустойчивость.
Пытаясь разобраться в этом непростом вопросе, ученые из Национальной лаборатории имени Лоуренса в Беркли (Lawrence Berkeley National Laboratory) с коллегами из научных учреждений США, Швейцарии и Испании экспериментально изучили свойства 10 энхансеров, участвующих в регуляции развития конечностей у мышиного эмбриона.
Все энхансеры, выбранные для анализа, удовлетворяют трем критериям. Во-первых, они эволюционно консервативны (то есть редко меняются в ходе эволюции), а значит, важны. Во-вторых, все они обеспечивают экспрессию подконтрольных генов в зачатках конечностей (рис. 1, левые колонки фотографий). В-третьих, все семь генов, чья экспрессия регулируется этими энхансерами (Sox5, Sox9, Shox2, Gli3, Tbx3, Fgf10, Sall1), необходимы для нормального развития конечностей. Мутации в этих генах ведут к разнообразным врожденным дефектам. Например, потеря гена Gli3 ведет к полидактилии, а отключение Fgf10 — к полному отсутствию лап у мышей.
Исследователи поочередно удаляли из геномов мышей эти 10 энхансеров при помощи CRISPR-Cas9 и смотрели, как будут развиваться конечности у эмбрионов. К немалому удивлению ученых, во всех 10 случаях конечности получились абсолютно нормальные! (рис. 1, правые колонки фотографий). Паттерны экспрессии подконтрольных генов тоже практически не изменились. Получается, что без любого из 10 энхансеров можно обойтись, что вообще-то очень странно, учитывая эволюционную консервативность этих регуляторных элементов.
Эти результаты свидетельствуют о дублировании функций. По-видимому, у каждого из 10 энхансеров есть «дублеры» — другие энхансеры с той же функцией. Чтобы проверить эту гипотезу, исследователи стали удалять энхансеры не по одному, а парами. Среди 10 выбранных энхансеров есть три подходящие пары: два энхансера регулируют ген Gli3, два другие — Shox2, еще два — Sox9 (рис. 1).
В двух случаях из трех (для Gli3 и Shox2) одновременное удаление пары энхансеров привело к серьезным дефектам. Потеря двух регуляторов гена Gli3 привела к снижению экспрессии этого гена, что на фенотипическом уровне проявилось в раздвоении больших пальцев на передних лапах. Удаление двух энхансеров гена Shox2 тоже привело к снижению экспрессии этого гена, а на уровне фенотипа — к недоразвитию бедренных костей.
Таким образом, хотя каждый энхансер по отдельности не является жизненно необходимым, удаление сразу двух энхансеров, регулирующих один ген, ведет к грубым нарушениям развития. Это согласуется с идеей о том, что «избыточные» энхансеры дублируют и подстраховывают друг друга.
Но авторы не остановились на достигнутом и провели еще одну серию экспериментов, чтобы выяснить, помогает ли подобная «подстраховка» в решении еще каких-нибудь проблем помимо утраты или несрабатывания одного из энхансеров. Для этого были выведены линии мышей с разными комбинациями работающих и неработающих энхансеров генов Gli3 и Shox2, у которых вдобавок был сломан один из этих генов на одной из двух гомологичных хромосом. Иными словами, это были мыши, гетерозиготные по отсутствию Gli3 (или Shox2), и при этом с неполными наборами энхансеров. Идея была в том, что если у мыши вместо двух нормально работающих копий гена есть только одна, то уровень экспрессии этого гена окажется в целом понижен, а фенотип станет более «уязвимым», то есть чувствительным к потере энхансеров. И такой мыши будет уже не всё равно, имеется ли у нее только один из двух энхансеров-дублеров или оба сразу.
Результаты в целом подтвердили эти ожидания (рис. 2). Например, у мышей с одной рабочей копией Gli3 и с двумя нормальными энхансерами большой палец лишь слегка раздваивается (рис. 2, вторая лапка сверху). Если в такой ситуации убрать любой из двух энхансеров, полидактилия становится более выраженной: получаются уже два отдельных больших пальца (третья и четвертая лапки). Аналогичные результаты получились и по гену Shox2.

Рис. 2. Лапы мышей с разными комбинациями испорченных и рабочих копий гена Gli3 и двух его энхансеров (mm1179 и hs1586). Сверху вниз снижается общий уровень экспрессии Gli3, а лапа становится всё более странной. Изображение из обсуждаемой статьи в Nature
По-видимому, каждый из «избыточных» энхансеров по отдельности вносит свой вклад в поддержание необходимого уровня экспрессии. Если все остальные системы работают нормально, то для формирования правильного фенотипа достаточно любого из двух энхансеров. Но если что-то пошло не так (например, забарахлила одна из двух копий гена), то индивидуальный вклад каждого энхансера становится заметен, и энхансеры перестают быть «избыточными».
Насколько широко распространено у мышей такое дублирование функций энхансеров? Точного ответа пока нет, потому что далеко не все мышиные энхансеры найдены или, тем более, экспериментально изучены. Однако имеющиеся данные позволяют находить возможные энхансеры и предсказывать некоторые их свойства. Для этого разработаны специальные методы. Вероятные энхансеры ищут по комбинации признаков, включая характерные последовательности нуклеотидов (к которым могут прикрепиться те или иные факторы транскрипции), эпигенетические модификации гистонов и особенности структуры хроматина. Имеющиеся детальные данные по экспрессии генов в разных частях мышиного эмбриона на разных стадиях развития позволяют судить о возможных функциях «предсказанных» энхансеров.
Проанализировав эти данные, авторы пришли к заключению, что, во-первых, множественные энхансеры характерны для большинства генов, управляющих развитием эмбриона, во-вторых, дублирование функций с большой вероятностью характерно для значительной доли энхансеров, регулирующих экспрессию этих генов.
По-видимому, помехоустойчивость развития чрезвычайно важна для млекопитающих (а может, и вообще для всех многоклеточных), и поэтому дополнительные регуляторные элементы, кажущиеся на первый взгляд избыточными, поддерживаются отбором, что и объясняет их эволюционную консервативность.
Источник: elementy.ru
 Опубликовано в рубрике
Опубликовано в рубрике