
 23 июня, 2018
23 июня, 2018 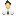 admin
admin 

Рис. 1. Результаты отбора на высокую и низкую успешность самцов дрозофил в конкуренции за самок. Черные точки соответствуют самцам из линий, отбиравшихся на максимальную успешность, белые — на неуспешность, серые — контрольные самцы из линий, не подвергавшихся искусственному отбору. A — успех в конкуренции за самку, B — выживаемость потомства (доля отложенных яиц, из которых благополучно развилась взрослая муха). Inbred — инбредные самцы, полученные от скрещивания брата с сестрой; Outbred — аутбредные самцы, чьи родители не состояли в близком родстве. Снижение показателей приспособленности у инбредных особей по сравнению с аутбредными отражает величину мутационного груза. Видно, что отбор на успешность вычистил из генофонда мутации, снижающие выживаемость потомства, но не справился с мутациями, снижающими брачный успех. Рисунок из обсуждаемой статьи в Science Advances
Одним из нерешенных вопросов теории полового отбора считается так называемый «парадокс токовища» (lek paradox): если самки всегда выбирают самцов по одним и тем же признакам, то почему в популяции сохраняется изменчивость по этим признакам, и почему все самцы не становятся одинаково привлекательными? Чтобы разобраться в этом, австралийские биологи изучили последствия искусственного отбора самцов дрозофил на повышенную и, наоборот, пониженную успешность в конкуренции за самок. Полученные результаты согласуются с гипотезой о том, что успешность самцов зависит от множества генов с малыми эффектами, и поэтому даже сильный отбор не успевает вычистить все возникающие в этих генах вредные мутации. Не получила подтверждений альтернативная гипотеза, согласно которой сохранение изменчивости по признакам, повышающим успешность самца, объясняется тем, что эти признаки сопряжены с вредными побочными эффектами.
«Элементы» уже рассказывали о двояком влиянии полового отбора на эволюцию видов. С одной стороны, женская привередливость при выборе партнера и конкуренция между самцами способствуют отбраковке вредных мутаций и снижению генетического груза (см.: Половой отбор защищает от вымирания, «Элементы», 23.05.2015). С другой стороны, результатом интенсивного полового отбора может стать развитие «дорогих» адаптаций, повышающих привлекательность самца (и его успешность в конкуренции за самок) ценой ухудшения каких-то других жизненно важных характеристик (см.: Половой отбор может довести до вымирания, «Элементы», 16.04.2018).
С этой двойственностью напрямую связаны два возможных механизма, которые теоретически могут объяснить «парадокс токовища» или «парадокс токового поведения» (lek paradox). «Парадокс» формулируется примерно так: если самки из поколения в поколение упорно выбирают самцов, обладающих определенными «привлекательными» признаками, то почему все самцы давным-давно не стали одинаково привлекательными (а женская избирательность не потеряла смысл)? Или, если говорить более общими словами, почему в популяциях сохраняется наследственная изменчивость по признакам, подвергающихся интенсивному однонаправленному отбору?
Первое объяснение предполагает, что привлекательность (или успешность) самца определяется его общим физическим состоянием, здоровьем, силой, энергичностью, то есть такими характеристиками, которые зависят от множества генов и потому являются хорошими «индикаторами приспособленности». Поскольку генов, влияющих на привлекательность, очень много, даже сильный отбор не может полностью вычистить из генофонда все возникающие в них негативные мутации. Поэтому наследственная изменчивость по привлекательности всегда сохраняется, а женская привередливость остается полезной как для самки (помогая ей выбрать партнера с лучшими генами), так и для популяции (способствуя уменьшению генетического груза).
Второе объяснение связано с вредными побочными эффектами признаков, повышающих успешность самца. Эти вредные эффекты могут затрагивать либо самих самцов (хороший пример рассмотрен в новости Почему половой отбор не может обеспечить всех баранов большими рогами, «Элементы», 07.10.2013), либо самок (см.: Самки медленнее стареют, если за них не конкурируют, «Элементы», 25.01.2014). В таком случае наследственная изменчивость по привлекательности может сохраняться благодаря балансирующему отбору.
Чтобы сравнить вклад этих двух механизмов, австралийские биологи провели эволюционный эксперимент на дрозофилах. Они решили посмотреть, что будет, если в одних подопытных линиях усилить отбор на привлекательность (или успешность) самцов, а в других вести отбор в противоположную сторону, позволяя размножаться только наименее привлекательным самцам. Идея была в следующем. Если верно первое объяснение, то усиленный отбор на успешность не приведет к негативным побочным эффектам и будет способствовать уменьшению генетического груза. Селекция неудачников, напротив, должна ухудшить генетическое здоровье популяции. Если же верно второе объяснение, то отбор на успешность, скорее всего, приведет к негативным побочным эффектам, а отбор неудачников освободит мух от ограничений, накладываемых половым отбором, и у них, может быть, какие-то компоненты приспособленности даже улучшатся.
Исходная лабораторная популяция мух Drosophila melanogaster была произведена от диких мух, пойманных в Квинсленде (Австралия) в 2012 году. Самцов разделили на «успешных» и «неудачников», сажая их по двое в пробирку с самкой. Самец-победитель, благополучно спарившийся с самкой, считался успешным, а его незадачливый конкурент, соответственно, объявлялся неудачником. Если за 90 минут ни один из самцов не добивался благосклонности самки, оба самца исключались из участия в эксперименте.
Успешные самцы стали родоначальниками четырех линий мух, в которых отбор на успешность продолжался в течение 14 поколений (success-selected lines). В этих линиях позволяли размножаться только тем самцам, которые победили другого самца (из той же линии) в состязании за самку. Самцы-неудачники стали прародителями четырех других линий, которые в течение 14 поколений подвергались отбору на неуспешность (failure-selected lines). Самцов точно так же заставляли попарно соревноваться за самку, но только победителей выкидывали, а проигравших оставляли. В эксперименте также участвовали четыре контрольные линии, произведенные от той же исходной популяции и жившие в таких же условиях, но не подвергавшиеся искусственному отбору.
По окончании селекции самцов из разных линий протестировали на успешность в конкуренции за самок. Теперь они должны были соревноваться с самцом из исходной популяции. Выяснилось, что селекция была результативной. В линиях победителей успешность самцов повысилась, а в линиях неудачников — снизилась по сравнению с контролем (рис. 1, А, три левые точки).
Авторы гордо сообщают, что им впервые удалось провести результативную селекцию по данному признаку. При этом они ссылаются на две недавние работы с описанием безуспешных попыток получить эволюционный ответ на такой отбор. Остается загадкой, почему при этом не упомянут намного более продолжительный и масштабный эксперимент Л. З. Кайданова, в котором самцы дрозофил отбирались на пониженную (или повышенную) половую активность (см.: О. В. Иовлева, 2016. Эксперимент длиною в полвека). Разница лишь в том, что Кайданов не заставлял самцов конкурировать друг с другом, а сажал одного самца к нескольким самкам и отбирал тех, кому требовалось больше (или меньше) времени, чтобы с кем-нибудь спариться. По-видимому, авторы обсуждаемой статьи просто не знают о работах Кайданова, которые во многом, мягко говоря, предвосхитили полученные ими выводы.
Чтобы оценить величину генетического груза (отягощенность геномов вредными мутациями), авторы применили стандартную методику, основанную на том, что при близкородственных скрещиваниях рецессивные вредные мутации переходят в гомозиготное состояние и проявляются в фенотипе. Соответственно, если инбридинг ведет к сильному снижению приспособленности, это говорит о большом генетическом грузе (такая же методика применялась в эксперименте на жуках, о котором рассказано в новости Половой отбор защищает от вымирания).
Оказалось, что инбридинг по-разному сказывается на разных аспектах приспособленности в разных подопытных линиях. Брачный успех самцов одинаково сильно снижается от инбридинга в линиях «победителей», «неудачников» и у контрольных мух (рис. 1, А). Это значит, что отбор не оказал заметного влияния на эффективность отбраковки рецессивных мутаций, снижающих брачный успех самцов. Данный результат согласуется с идеей о том, что успешность самца зависит от огромного количества генов с малыми эффектами. Этих генов так много, что все возникающие в них вредные мутации никакой отбор не в состоянии вычистить. Поэтому наследственная изменчивость по успешности (или привлекательности) самцов всегда будет сохраняться, а привередливость самок не станет излишней.
С другой стороны, интенсивный отбор на успешность, кажется, помог вычистить вредные мутации, снижающие выживаемость потомства. Это проявляется в том, что выживаемость потомства инбредных самцов из линий «победителей» не ниже, чем у аутбредных самцов из тех же линий (правая черная точка на рис. 1, B расположена даже выше левой). Этим «победители» отличаются от контрольных мух и «неудачников», у которых инбредная депрессия выживаемости хорошо выражена (правые белая и серая точки на рис. 1, B расположены ниже левых).
Можно было бы ожидать, что у «неудачников» генетический груз увеличится по сравнению и с «победителями», и с контрольными мухами. Это было четко показано в эксперименте Кайданова, но в обсуждаемой работе этого не произошло — скорее всего, просто потому, что селекция продолжалась недостаточно долго.
В поисках возможных негативных эффектов усиленного отбора на привлекательность авторы проверили мух по трем признакам: устойчивость к обезвоживанию, плодовитость самок и успешность самцов в конкуренции спермы (в последнем случае брали самок, уже спарившихся с другим самцом, и позволяли им спариться повторно с тестируемым самцом, а потом смотрели, отцом скольких потомков окажется тестируемый самец). Ни по одному из трех признаков мухи из линий «победителей» не уступили контрольным, а по третьему даже заметно превзошли их.
Таким образом, никаких негативных эффектов отбора на успешность обнаружить не удалось. Зато было найдено целых два положительных эффекта: уменьшение груза вредных мутаций, влияющих на выживаемость потомства, и рост успешности в конкуренции спермы. Всё это согласуется с первым из двух упомянутых объяснений «парадокса токовища», но не со вторым.
Исследование дало новые аргументы в пользу нескольких важных идей, которые, несмотря на хорошую теоретическую обоснованность, по-прежнему нуждаются в эмпирической поддержке:
1) половой отбор помогает избавляться от генетического груза;
2) на успех самца в конкуренции за самок влияет большое число генов с малыми эффектами;
3) даже сильный отбор на успешность не ведет к исчезновению наследственной изменчивости по этому признаку, потому что признак зависит от слишком большого количества генов;
4) привередливость самок при выборе партнера остается полезным признаком, потому что более привлекательные (или конкурентоспособные) самцы являются носителями лучших генотипов, и дети от них будут более приспособленными.
К недостаткам работы следует отнести, помимо необъяснимого игнорирования работ Кайданова, недостаточно четкое разграничение привлекательности, конкурентоспособности и половой активности самцов. Авторы полагают, что, сажая самку к двум самцам, они давали ей возможность выбора, и поэтому то, с кем из двоих она спарится, зависело от ее предпочтений и от привлекательности самца. Однако самцы дрозофил в подобных ситуациях не только соревнуются в искусстве соблазнения, но и отталкивают соперника, так что победить в итоге может не тот, кто больше понравился самке, а тот, кто агрессивнее, сильнее или наглее. Дело это тонкое. Трудно сказать, на что в действительности шел отбор — на привлекательность самца (как полагают авторы) или на его половую активность (как в ходе своего эксперимента полагал Кайданов). Впрочем, на основные выводы работы это мало влияет.
Источник: elementy.ru
 Опубликовано в рубрике
Опубликовано в рубрике