
 3 апреля, 2018
3 апреля, 2018 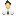 admin
admin 

Рис. 1. Схема строения и общий вид фага лямбда. Подписаны названия белков (C, D, E, W, B’, FII, U, V, tfa, stf, M, L, J, H); в скобках указано количество молекул данного белка в вирусной частице. За связывание с рецептором — поверхностным белком жертвы (бактерии Escherichia coli) — отвечает белок J, находящийся на конце ножки вируса. Изображение из статьи S. V. Rajagopala et al., 2011. The protein interaction map of bacteriophage lambda
Американские биологи расшифровали эволюционный механизм, позволяющий бактериофагу лямбда в лабораторных экспериментах вырабатывать новый способ заражения бактериальных клеток. Эволюционное новшество возникает в ответ на приобретение бактериями устойчивости к старому способу вирусной атаки, основанному на прикреплении вирусного белка J к бактериальному поверхностному белку LamB. В этой ситуации отбор сначала закрепляет мутации гена J, усиливающие старую функцию. «Платой» за лучшее связывание с LamB становится дестабилизация белка J. В итоге получаются вирусы, у которых при одном и том же геноме белок J может иметь две разные пространственные конфигурации. При этом вирусные частицы с «правильно» свернутым J заражают жертв старым способом, а другие (с таким же геномом, но с «неправильно» свернутым J) делают это по-новому, прикрепляясь к другому поверхностному белку бактерии (OmpF). В дальнейшем оба варианта могут стабилизироваться путем закрепления дополнительных мутаций, что фактически приводит к разделению исходного вируса на два вида. Работа подтверждает старую, но имеющую мало экспериментальных подтверждений идею о том, что новые функции могут развиваться через промежуточный этап дестабилизации фенотипа с последующей стабилизацией («генетической ассимиляцией») удачных ненаследственных отклонений.
Теоретики давно обсуждают возможную роль дестабилизации фенотипа в появлении эволюционных новшеств. Предполагается, что новшества могут возникать по следующей схеме: «исходный стабильный фенотип → дестабилизирующее воздействие (например, резкое изменение среды) → рост негенетической изменчивости → генетическая ассимиляция удачного фенотипа, то есть закрепление мутаций, стабилизирующих тот вариант фенотипа, который оказался адаптивным в новых условиях». Об этой модели подробно рассказано в новости Дестабилизация развития — путь к эволюционным новшествам («Элементы», 13.07.2009). Идея выглядит логичной, однако прямых экспериментальных подтверждений у нее пока немного (см. ссылки в конце новости).
Американские биологи, работающие с бактериофагом лямбда (см. Lambda phage), пополнили коллекцию подтвержденных примеров действенности данного механизма еще одним экспонатом.
Изучалось эволюционное новшество, систематически возникающее у фага λ в определенных условиях. В норме этот фаг заражает своих жертв, кишечных палочек Escherichia coli, прикрепляясь к поверхностному белку (рецептору) LamB. Однако жертвы могут выработать устойчивость к вирусу путем накопления мутаций, снижающих уровень экспрессии этого рецептора. Количество молекул LamB на поверхности бактериальных клеток уменьшается, и вирусу становится не за что ухватиться.
В присутствии устойчивых бактерий вирусы подвергаются интенсивному отбору на способность как можно эффективнее цепляться за те немногие молекулы LamB, которые еще остались. Отбор последовательно закрепляет 4–7 мутаций в вирусном гене, кодирующем белок J. Этот белок располагается на конце ножки вируса и отвечает за прикрепление к LamB (рис. 1).
Неожиданным образом эти мутации не только повышают прочность связи с LamB, но и придают белку J новую способность — прикрепляться к другому поверхностному белку бактерии, OmpF. Фактически вирус вырабатывает новый способ заражения бактериальных клеток. Такие вирусы успешно заражают бактерий, даже вовсе лишенных LamB (J. R. Meyer et al., 2012. Repeatability and Contingency in the Evolution of a Key Innovation in Phage Lambda).
Недавно было показано, что получившиеся вирусы-генералисты (способные заражать своих жертв двумя способами) могут затем снова специализироваться, то есть утратить один из двух способов заражения, оптимизировав другой. Это может привести к видообразованию (разделению на два вида). Хотя у фагов λ нет полового размножения, у них есть рекомбинация — обмен участками генома между вирусами, заразившими одну и ту же клетку. С точки зрения эволюционных последствий это почти то же самое, что и половое размножение. Оказалось, что адаптация к хозяевам, у которых есть только один из двух рецепторов (LamB или OmpF), ведет к разделению вирусов-генералистов на две специализированные группы, каждая из которых способна заражать только один тип жертв. Такие вирусы уже не могут меняться друг с другом генами, потому что в их геномах закрепляются несовместимые мутации (J. R. Meyer et al., 2016. Ecological speciation of bacteriophage lambda in allopatry and sympatry). Поэтому их вполне можно считать разными видами.
В ходе нового исследования, результаты которого опубликованы в журнале Science, американские вирусологи разобрались в том, каким образом белку J вирусов-генералистов удается совмещать две функции. У клеточных организмов такие эволюционные изменения обычно происходят за счет дупликации гена с последующим разделением функций между копиями. Но у фагов-генералистов ген J не дуплицирован.
Альтернативный механизм связан с дестабилизацией белка. Авторы предположили, что мутации, закрепившиеся в гене J у фагов-генералистов, внесли элемент хаоса в процесс сворачивания кодируемого белка (см. Фолдинг белка). Возможно, белок J у фагов-генералистов может принимать две разные конформации, одна из которых связывается с LamB, а другая — с OmpF.
Дестабилизация пространственной структуры белка часто сопровождается снижением его устойчивости к повышению температуры. Поэтому проверку своей гипотезы ученые начали с оценки выносливости вирусов к перегреву. Для этого они в течение часа выдерживали вирусные частицы с разными генотипами при разных температурах (от 37°С — оптимальной температуры для фага λ до смертельных 55°С) и смотрели, какой процент вирусов сохранит жизнеспособность. В эксперименте использовались генотипы, соответствующие разным этапам изученного ранее эволюционного пути от исходного вируса (связывающегося только с LamB) к вирусу-генералисту. Изучаемые вирусы различались только мутациями в гене J, а весь остальной геном у них был одинаковый.
Результаты подтвердили ожидания исследователей (рис. 2). Выяснилось, что мутации в гене J, которые в ходе адаптации вирусов к жертвам с пониженной экспрессией LamB повышали сродство J к LamB, а затем дали возможность связываться также и с OmpF, попутно снижали термостабильность белка J.

Рис. 2. Термоустойчивость вирусных частиц с разными генотипами. По вертикальной оси — доля вирусных частиц, сохранивших жизнеспособность после часа выдерживания при температуре, указанной на горизонтальной оси. Черная сплошная линия — «дикий тип» (0 мутаций), то есть вирус с исходной версией белка J, прикрепляющийся только к рецептору LamB. Черная пунктирная линия — 3 мутации в белке J, повышающие сродство к LamB, но еще не дающие возможности связываться с OmpF. Зеленые линии — генотипы с 4–7 мутациями в J, способные связываться с обоими рецепторами (вирусы-генералисты). Рисунок из обсуждаемой статьи в Science
Впрочем, это само по себе еще ни о чем говорит, потому что мутации, меняющие аминокислотные последовательности белков, часто снижают устойчивость белков к перегреву — это дело обычное. Но в данном случае всё оказалась интереснее. Авторы проверили термочувствительность упомянутых выше вирусов — потомков генералистов, которые в ходе дальнейшей эволюции снова стали специалистами, утратив способность прикрепляться к одному из двух рецепторов. У этих вирусов — «вторичных специалистов» в гене J еще больше мутаций по сравнению с исходным вариантом, чем у генералистов. Однако устойчивость к нагреванию у них оказалась такой же высокой, как и у «диких» вирусов. Таким образом, «генерализующие» мутации снизили термостабильность, а «специализирующие» снова ее повысили.
Самый интересный результат был получен, когда авторы проанализировали временную динамику разрушения вирусных частиц при оптимальной для них температуре 37°С. В опыте использовали штаммы с максимальной и минимальной термоустойчивостью, то есть вирусы «дикого типа» и генералистов с семью мутациями (этим двум генотипам соответствуют черная и зеленая сплошные линии на рис. 2). Оказалось, что генералисты со временем разрушаются быстрее, чем «дикие» вирусы. Это ожидаемый результат, потому что от менее термоустойчивых вирусов следует ожидать и меньшей устойчивости при оптимальной температуре. Интереснее другое: исследователи обнаружили, что в течение первых двух суток разрушение вирусов дикого типа идет с постоянной скоростью, тогда как генералисты намного быстрее деградируют в первые сутки, чем во вторые (рис. 3).

Рис. 3. Динамика разрушения вирусных частиц при 37°С. Черные линии — вирусы с исходным генотипом, зеленые — генералисты с семью мутациями в гене J. Видно, что первые разрушаются с постоянной скоростью, а вторые быстрее разрушаются в первый день, чем во второй. Это говорит о разнородности вирусов-генералистов (среди них есть устойчивые и неустойчивые индивиды), хотя генотип у них одинаковый. Рисунок из обсуждаемой статьи в Science
Но если все вирусные частицы в выборке одинаковы, то они должны разрушаться с постоянной скоростью. Полученный результат говорит о том, то вирусы-генералисты, по-видимому, представлены двумя разными фенотипами, один из которых неустойчив и разрушается быстро (в основном в течение первого дня), а другой — медленно (примерно с той же скоростью, что и «дикие» вирусы). Однако генотип у всех генералистов один и тот же. Стало быть, речь идет о негенетической изменчивости. Скорее всего, дело тут в разных вариантах сворачивания белка J.
Дальнейшие эксперименты дали ряд косвенных подтверждений этому предположению. Удалось показать, что быстро разрушающаяся фракция вирусов-генералистов преимущественно связывается с OmpF, а медленно разрушающаяся — с LamB. Это видно, например, из того, что по мере разрушения вирусных частиц среди вирусов-генералистов остается все меньше способных связываться с OmpF, в то время как доля связывающихся с LamB растет. А если отобрать те вирусы, которые связались с OmpF, то оставшиеся вирусы, во-первых, лучше связываются с LamB, чем исходная смесь, во-вторых, со временем разрушаются медленнее. Дополнительные эксперименты подтвердили, что изменчивость у вирусов-генералистов действительно негенетическая, то есть ненаследственная.
В итоге вырисовалась следующая схема появления эволюционного новшества. В ходе адаптации к жертвам с пониженной экспрессией рецептора LamB отбор стал поддерживать у вирусов такие мутации в гене J, которые позволяли белку J прочнее связываться с LamB. Это достигалось ценой дестабилизации белка J, который в результате стал иногда сворачиваться неправильно. После приобретения четырех таких мутаций (каждая из которых повышала сродство J к LamB) у белка J появился новый вариант сворачивания, который позволял связываться с другим рецептором — OmpF. Белок с новой функцией возник как один из вариантов фенотипа в рамках негенетической изменчивости. При одном и том же геноме часть вирусов теперь могла связываться с OmpF, в то время как другие обладатели того же генотипа связывались с LamB. Так появились вирусы-генералисты. При этом каждая отдельная вирусная частица намного эффективнее связывалась с одним из двух рецепторов, чем с другим. Генерализация произошла на уровне популяции, а не индивида.
В дальнейшем вирусы-генералисты могут снова специализироваться, попав в подходящие условия (то есть получив доступ к жертвам, у которых есть либо только LamB, либо только OmpF). В ходе специализации закрепляются мутации, повышающие вероятность того, что белок J свернется выгодным в данной ситуации образом. В результате белок J снова стабилизируется, то есть начинает сворачиваться только одним способом. При этом восстанавливается также и устойчивость белка (и всего вируса) к повышенной температуре.
Таким образом, найден яркий пример появления новой функции через промежуточный этап, связанный с дестабилизацией фенотипа. Остается неясным, как часто возникают таким способом эволюционные инновации у вирусов и клеточных организмов. Ответ на этот вопрос должны дать дальнейшие исследования.
Источник: elementy.ru
 Опубликовано в рубрике
Опубликовано в рубрике