
 28 февраля, 2018
28 февраля, 2018 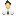 admin
admin 


Рис. 1. Три окрасочные морфы палочников Timema cristinae: зеленая полосатая (green striped), зеленая бесполосая (green unstriped) и темная или меланистическая (melanistic). Первый вариант окраски лучше маскирует насекомое на листьях чамиза (Adenostoma fasciculatum), второй — на листьях краснокоренника (Ceanotus spinosus), третий — на стеблях обоих растений. Фото — из популярного синопсиса к обсуждаемой статье в Science, рисунок — из статьи A. A. Comeault et al., 2015. Selection on a Genetic Polymorphism Counteracts Ecological Speciation in a Stick Insect
Двадцатипятилетние наблюдения за палочниками Timema cristinae в Калифорнии позволили оценить предсказуемость эволюции на примере двух параметров окраски: цвета и рисунка. Оказалось, что рисунок (наличие или отсутствие полосок) эволюционирует предсказуемо: соотношение полосатых и бесполосых форм можно предсказать на несколько лет вперед, зная, как оно менялось в прошлые годы. Главный фактор, определяющий направленность отбора, в данном случае всего один, и он лежит на поверхности: это относительная частота встречаемости форм (редкая форма получает преимущество, что порождает регулярные, предсказуемые колебания). Напротив, соотношение палочников, различающихся по цвету (зеленых и темно-серых), определяется множеством слабоизученных факторов и потому не может быть предсказано на основе имеющихся данных. По-видимому, микроэволюционные процессы в принципе можно предсказывать довольно точно, но для этого необходимо глубокое понимание всех причин, влияющих на направленность отбора, а их может быть много.
Вопрос о предсказуемости эволюции издавна волнует биологов. С одной стороны, известно, что случайные события играют в эволюции важную роль. Это и случайный во многих отношениях мутагенез (см.: Ранние этапы адаптации предсказуемы, поздние — случайны, «Элементы», 03.03.2015; Предсказуемый рост приспособленности достигается непредсказуемыми путями, «Элементы», 30.06.2014), и генетический дрейф (см.: Эволюционный эксперимент на ящерицах показал действенность «эффекта основателя», «Элементы», 09.03.2012), и колебания условий среды, которые могут резко менять направленность отбора (см.: Расшифрованы генетические основы быстрых эволюционных изменений размера клюва у дарвиновых вьюрков, «Элементы», 25.04.2016), и всевозможные редкие, маловероятные события вроде случайного попадания какого-нибудь животного на уединенный остров (см.: Новый вид дарвиновых вьюрков возник на глазах у исследователей, «Элементы», 04.12.2017).
Подобные случайности могут иметь далеко идущие эволюционные последствия. Поэтому на практике, как правило, предсказать дальнейший ход эволюции по данным о прошлых событиях оказывается крайне трудно или вообще невозможно, а эволюционной биологии приходится довольствоваться статусом «описательной», а не «предсказательной» науки (Peter R. Grant, B. Rosemary Grant, 2002. Unpredictable Evolution in a 30-Year Study of Darwin’s Finches).
С другой стороны, естественный отбор — процесс вовсе не случайный, а наоборот, направленный и детерминистский. К тому же многочисленные ограничения самой разной природы (молекулярные, онтогенетические, экологические) резко сужают круг доступных эволюционных траекторий (см.: Пути эволюции предопределены на молекулярном уровне, «Элементы», 12.04.2006; Эволюция белков сдерживается низкой проходимостью ландшафта приспособленности, «Элементы», 09.02.2015). Поэтому в какой-то степени эволюция всё же должна быть предсказуемой, что было подтверждено в ряде работ (см.: Способы адаптации бактерий к разным температурам оказались предсказуемыми, «Элементы», 16.03.2017; M. Lässig et al., 2017. Predicting evolution). На частичную предсказуемость и повторяемость эволюции недвусмысленно намекает широкая распространенность параллелизмов, то есть независимого возникновения сходных признаков и жизненных форм в разных эволюционных линиях (см., например: Параллельная эволюция изучена в эксперименте на бактериях, «Элементы», 01.02.2012; Параллелизмы — результат быстрой эволюции сенсорных рецепторов, «Элементы», 20.09.2011; Цихлиды — живая модель независимой параллельной эволюции, «Элементы», 14.11.2012; Параллельная эволюция эусоциальности у пчел: принцип общий, эволюционные маршруты — разные, «Элементы», 15.05.2015).
Биологи из США, Великобритании и Канады предприняли очередную попытку оценить предсказуемость эволюции на примере нелетающих палочников Timema cristinae — обитателей калифорнийского чапараля. Об этих насекомых и о том, почему на них удобно изучать микроэволюцию, рассказано в новости Анализ геномов калифорнийских палочников помог оценить соотношение случайности и закономерности в эволюции, «Элементы», 20.05.2014.
Направленность отбора, действующего на палочников, во многом определяется птицами, которые избирательно выедают особей, более заметных на окружающем фоне. Заметность, в свою очередь, зависит от окраски палочника и от того, на каком растении он сидит. У T. cristinae есть три дискретных варианта окраски, или окрасочных морфы (рис. 1):
1) зеленая полосатая, выигрышная на узких листьях растения Adenostoma fasciculatum (чамиз),
2) зеленая бесполосая, маскирующая насекомое на широких листьях растения Ceanothus spinosus (краснокоренник),
3) темно-серая (меланистическая), помогающая прятаться на стеблях обоих растений.
Зеленые полосатые палочники преобладают на аденостоме (но встречаются и на краснокореннике), зеленые неполосатые доминируют на краснокореннике (но присутствуют и на аденостоме), а темных палочников в среднем по 10% на обоих растениях.
Изменчивость по окраске определяется комбинацией двух признаков: цвета (зеленого или темно-серого) и рисунка (наличия или отсутствия полосок), причем второй признак проявляется только у зеленых индивидов. Оба признака имеют высокую наследуемость, то есть зависят в основном от генов, а не от среды. Ранее было показано, что зеленый цвет и отсутствие полосок — доминантные варианты, а темный цвет и полосатость — рецессивные.
В статье, опубликованной в свежем выпуске журнала Science, авторы описали результаты многолетнего мониторинга, проводившегося в горах близ Санта-Барбары (Калифорния, США) с 1990 по 2017 год. В общей сложности было учтено 34 383 палочника из 545 точек в пространстве и времени (545 комбинаций «год — точка на местности — растение-хозяин»).
Для начала ученые разобрались с генетическим базисом изучаемых признаков. Секвенирование геномов представителей разных окрасочных морф и полногеномный поиск ассоциаций позволили идентифицировать участок генома протяженностью в 10,5 млн пар оснований, от которого зависят сразу оба признака: и цвет, и рисунок. В пределах этого участка, получившего название «локус Mel-Stripe», ограничена мейотическая рекомбинация (см. Кроссинговер), то есть он наследуется как единое целое. Оказалось, что у палочников есть три варианта (аллеля) локуса Mel-Stripe. Авторы обозначили их буквами u (unstriped, бесполосый), s (striped, полосатый) и m (melanistic, меланистический). Аллель u доминирует над s и m, s доминирует над m. Таким образом, особи с генотипами uu, us и um — зеленые бесполосые, генотипы ss и sm соответствуют зеленому полосатому фенотипу, а обладатели генотипа mm имеют темно-серую окраску.
Следующая задача, которую авторы поставили перед собой, заключалась в том, чтобы убедиться, что частоты аллелей Mel-Stripe быстро меняются под действием отбора. Для этого были проанализированы генетические данные, собранные в ходе трех предыдущих исследований (одно из них, связанное с пересадкой палочников с одних растений на другие, описано в новости Анализ геномов калифорнийских палочников помог оценить соотношение случайности и закономерности в эволюции, «Элементы», 20.05.2014). Оказалось, что локус Mel-Stripe действительно склонен к ускоренной эволюции: частоты аллелей в этом локусе меняются быстрее, чем в других участках генома. Это трудно объяснить чем-либо, кроме интенсивного отбора, направленность которого может меняться в зависимости от условий.
Предсказуемость эволюции двух признаков (цвета и рисунка) оценивалась при помощи моделей авторегрессии — скользящего среднего (Autoregressive–moving-average model). Авторы рассчитали, с какой точностью можно предсказать изменения частоты встречаемости признака на несколько (3–10) лет вперед на основе данных о его частоте за несколько предшествующих лет.
Оказалось, что предсказуемость эволюции цвета (колебаний соотношения зеленых и темно-серых палочников) невелика: данные о прошлом объясняют лишь 14% последующих изменений. По-видимому, такая низкая предсказуемость обусловлена тем, что направленность отбора в данном случае зависит от множества малоизученных причин, не имеющих прямого отношения к предшествующей истории популяции. Например, ученые обнаружили, что можно увеличить предсказательную силу модели, включив в нее данные по весенним температурам. Как выяснилось, темно-серые палочники обычно становятся чуть более многочисленными в годы с теплой весной. Это логично: в такие годы растения подсыхают, и у них становится больше потемневших веточек, на которых темные палочники не так заметны, как зеленые. Впрочем, влияние температуры может быть двояким: дополнительные эксперименты показали, что темные палочники хуже зеленых переносят сильный перегрев. Таким образом, жаркая погода, с одной стороны, повышает долю темных палочников (через подсыхание растений и лучшую маскировку), с другой — снижает ее (через избирательную гибель темных палочников при сильном перегреве). Первый эффект, по-видимому, чуть сильнее второго, поэтому в суммарном зачете темные палочники оказываются в выигрыше в теплые годы. Но даже с учетом температуры точность предсказаний остается низкой.
О том, что направленность отбора по цвету определяется множеством причин, свидетельствует также и то, что темные палочники меньше подвержены грибковым инфекциям и успешнее конкурируют за половых партнеров, чем две другие морфы.
Эксперименты, предпринятые для оценки выживаемости шести генотипов на одном из кормовых растений (а именно на аденостоме, где преобладает зеленая полосатая морфа) дали противоречивые результаты: получилось, что в течение теплого сезона лучше всех выживают гомозиготы ss, а зиму лучше переносят гетерозиготы sm. Оба генотипа, как мы помним, дают один и тот же фенотип (зеленый полосатый). Эти результаты свидетельствуют о том, что локус Mel-Stripe влияет не только на окраску, а отбор по этому локусу определяется множеством малоизученных факторов. Этим, скорее всего, и объясняется низкая предсказуемость эволюции. Чтобы предсказывать ее точнее, нужно детально разобраться в сложном переплетении причинно-следственных связей, определяющих направленность отбора.
Предсказуемость эволюции рисунка, то есть соотношения полосатых и бесполосых зеленых палочников, оказалась намного выше. Предсказанные значения строго коррелируют с реальными (r2 = 0,86, то есть данные по предшествующим годам объясняют 86% будущих колебаний). Высокая предсказуемость объясняется тем, что эволюционная динамика в данном случае очень проста: доля полосатых насекомых регулярно колеблется вверх-вниз, и если в этом году она выросла, то в следующем наверняка упадет. Самым правдоподобным объяснением такой картины является частотно-зависимый балансирующий отбор (так называют ситуацию, когда выгодно быть обладателем редкого вариант признака). Видимо, дело в том, что птицы учатся находить более массовую добычу, и поэтому редкий вариант оказывается в выигрыше — но только до тех пор, пока он остается редким.
Чтобы проверить гипотезу о частотно-зависимом отборе, авторы провели дополнительный эксперимент. Десять тщательно очищенных от палочников кустов аденостомы были заселены зелеными полосатыми и зелеными бесполосыми палочниками в пропорции 4:1, а другие десять кустов — теми же морфами в пропорции 1:4. В общей сложности пересадили 1000 палочников, по 50 насекомых на растение. Через пять дней насекомых снова собрали и пересчитали, определив таким образом, сколько полосатых и бесполосых особей съели птицы на каждом растении. Оказалось, что на тех кустах, где преобладали бесполосые палочники, птицы ели в основном их, так что среди выживших насекомых доля полосатых резко выросла. Там же, где преобладали полосатые палочники, птицы поедали обе формы примерно в одинаковой пропорции, так что ни одна из форм не имела преимущества, и их соотношение не изменилось. Этот результат в целом соответствует идее о частотно-зависимом отборе: полосатые палочники имеют сильное преимущество, когда их 20%, и не имеют преимущества, когда их 80%. Не исключено, что бесполосые палочники могут даже иметь селективное преимущество на аденостоме (где вообще-то всегда доминируют полосатые), если их доля меньше 20%. Но это еще нужно проверить.
В заключительной части статьи авторы сравнивают свои данные с результатами других (пока весьма немногочисленных) многолетних проектов по изучению эволюции «в реальном времени», включая наблюдения супругов Грант за дарвиновыми вьюрками и мониторинг окрасочных морф березовой пяденицы (см.: Промышленный меланизм бабочек получил генетическое объяснение, «Элемент», 06.06.2016). Для уверенных выводов данных пока недостаточно, но складывается впечатление, что низкая предсказуемость микроэволюции (как в случае с цветом у палочников) — это скорее норма, а высокая предсказуемость (как в случае с рисунком) — скорее исключение. Впрочем, данные по палочникам, по мнению авторов, свидетельствуют о том, что главной причиной низкой предсказуемости является не какая-то фундаментальная, внутренне присущая микроэволюции хаотичность, а множественность факторов, влияющих на направленность отбора, и неполнота наших знаний о них. Чтобы точнее предсказывать эволюцию, нужно глубже вникать в детали.
Источник: elementy.ru
 Опубликовано в рубрике
Опубликовано в рубрике